En förutsättning för växtförädling är att det finns genetisk variation inom en art, det vill säga att olika växtindivider har olika genvarianter, alleler, och därmed olika egenskaper. En potatissort kan till exempel vara resistent mot en svampsjukdom, en annan potatissort saknar den egenskapen, men bär på en allel som gör att den är motståndskraftig mot ett visst virus. Genetiskt variation orsakas av att det uppstår förändringar, mutationer, i DNA. Mutationer kan uppstå spontant och är en förutsättning för evolutionen. De kan ge positiva, negativa eller neutrala effekter och påverkan på växtens förmågan att föröka sig. Ibland är mutation som negativ för en vild växt men positiv ur perspektivet att vi människor vill odla den.
Från vilda växter till odlade
För omkring 10 000 år sedan blev människan bofast och började odla växter och hålla lantbruksdjur. Sedan dess har de växtindivider med egenskaper som har varit till mest nytta för människan valts ut och förökats. Över tid har då vilda växter mer och mer anpassats efter människans behov. Det är alltså fråga om ett mänskligt och inte ett naturligt urval.
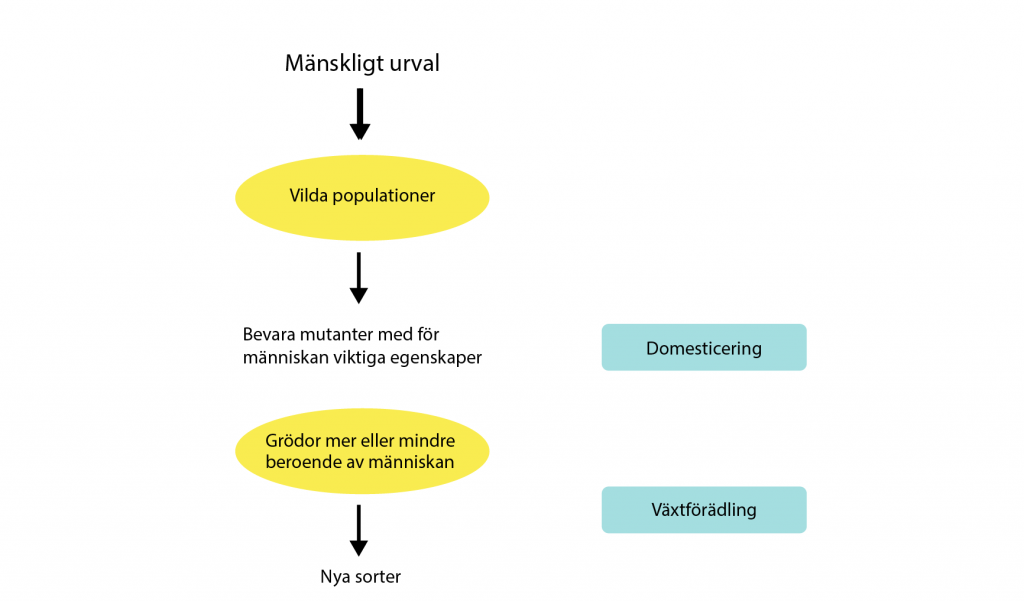
Denna så kallade domesticering innebar stora förändringar av de växter människan samlade in. Från att ha varit anpassade till sin livsmiljö i naturen anpassades de växterna till människans odlingar. Gynnsamt för människan – men inte en växt i naturen – är till exempel ett gräs som där dess frön inte faller till marken när det mognat (dråsar) utan i stället sitter fast på axet så att de kan skördas. Det här kan man tydligt se i skillnaden mellan den odlade majsen, där majskornen sitter kvar i majskolven, och den vilda majsen, teosint, som har få frön på glesa ax som när de mognar faller till marken. Det är en viktig egenskap för att den vilda majsen ska sprida sig effektivt.

Frön faller till marken och djur äter upp dem. Det hårda skalet gör att fröna går närmast oskadda förbi djurets mag-tarmkanal. Djuret vandrar iväg och via avföringen kommer teosintes frön i marken på annan plats än där den ursprungligen växte. Teosinten har spridit sina gener. Egenskaper som passar oss människor är många frön (majskorn), med mjukt skal som sitter fast på axet när de mognat och så ser dagens majs ut. Det beror på att vi människor över lång tid föredragit att odla majs med genetiska förändringar som givit den här typen av egenskaper.
Ett annat exempel är de olika typerna av kål. Blomkål, brysselkål, kålrabbi, vitkål och broccoli är alla samma art och härstammar från den vilda kålen (Brassica oleracea). Att det till exempel utvecklas ett blomkålshuvudet beror på mutationer i vildkålens blomningsgener. Det gör att den har svårigheter att blomma. Den försöker, men bildar istället ett blomkålshuvud. Ur kålens perspektiv är det negativt eftersom den inte kan sprida sina gener om den inte blommar, för människan är det positivt eftersom vi får mer att äta. Den typen av mutationer skulle snabbt ha försvunnit ur vildkålspopulationen om vi människor inte tagit hand om dem. Men att det rörde sig om genvarianter och mutationer hade människan ingen aning om under de tusentals år hon avvände urvalsmetoden vid odling. Urvalsmetoden dominerande sättet som nya varianter av grödor togs fram på långt in på 1900-talet.
Genetikens fader – munken Gregor Mendel
Munken Gregor Mendel var den vetenskapliga bakgrunden till ärftlighet på spåret när han korsade ärtor i klostret i nuvarande Brno i Tjeckien. Han publicerade en artikel 1865 med titeln Försök med växtbastarder, men ingen intresserade sig för hans forskning. Mendel skickade ett exemplar av artikeln till Charles Darwin som publicerat Om arternas uppkomst några år tidigare. Men vid Darwins död stod artikeln oläst i en bokhylla. Vid sekelskiftet, 16 år efter Mendels död, återupptäcktes hans forskning. Det blev startskottet till en mer systematisk växtförädling och Mendel kallas idag genetikens fader. Termerna homozygot och heterozygot introducerades 1902 och strax därefter termen genetik.
Traditionell mutationsförädling
Under 1920-talet upptäckte forskare att den genetisk variationen i ett förädlingsmaterial kunde mångfaldigas om det utsattes för strålning. På 1940-talet började även mutagena ämnen att användas för att öka den genetiska variationen. Mutationsförädling, där strålning eller mutagena ämnen används, var särskilt populärt under 1950- och 1960-talet, inte minst i Sverige. Under 1980-talet började intresset för mutationsförädling att avta, men vid millenniumskiftet introducerades en ny metod, TILLING, och i och med det ökade intresset igen. De traditionella mutagenesmetoderna är som ett skott i mörkret. De leder till slumpmässiga mutationer över hela växtens genom och tidigare var man tvungen att vänta till dess plantorna växt upp för att avgöra vilka växtmutanter som bar på intressanta egenskaper. TILLING-metoden innebär att man med hjälp av DNA-analyser på ett tidigt stadium kan välja ut intressanta mutanter. Därmed snabbades processen upp avsevärt. Idag är det oftast det mutagena ämnet etylmetansulfonat (EMS) som används, kombinerat med TILLING.
Växter som muterats med strålning eller mutagena ämnen leder enligt EU:s GMO-lagstiftning till en genetiskt modifierad organism (GMO), men undantas reglering. De behandlas alltså inte på något annat sätt än resultat efter en korsning mellan till exempel två vetesorter.
Hybridförädling
Vid hybridförädling korsas inavlade plantor. Att de är inavlade innebär att de är homozygota för de flesta av sina gener. De plantor som korsas ska vara så genetiskt olika varandra som möjligt. Resultatet av korsningen mellan dessa så kallade rena linjer blir starkt heterozygota hybridplantor som är mer produktiva än sina föräldrar. Detta kallas heterosiseffekten eller hybridvigör och det är hybriderna som odlas. Heterosisffekten kvarstår dock bara i en generation. Frön från en hybridplanta är sinsemellan väldigt olika genetiskt och om de används som utsäde blir resultatet av varierande kvalitet. Av den anledningen köper de flesta lantbrukare nytt utsäde varje år. Hybridförädling används på många grödor, till exempel majs och sockerbeta.
Artkorsningar och polyploidi
Under tidigt 1930-tal upptäcktes att polyploidi spelat en viktig roll i växternas evolution och att många viktiga grödor, som potatis och vete var polyploida. Intresset väcktes då för artificiellt framtagna polyploider. Mot slutet av 1930-talet publicerades en artikel där forskare visade att kromosomtalet kunde fördubblas om växtmaterial utsattes för det toxiska ämnet colchicin. Colchicin finns i växten tidlösa och påverkar celldelningen genom att hämma bildandet av fungerande mikrotubuli. Mikrotubulis uppgift under celldelningen är att se till att de två celler som blir resultatet av en delning får lika många kromosomer.
Artificell polyploidisering används bland annat i samband med att olika arter korsas. Ett exempel är rågvete. Råg och vete är två olika arter och därför svåra att korsa. Det embryo som utvecklas vid en korsning har svårt att överleva utan människans assistans. Embryot måste därför dissekeras fram och odlas på ett näringsmedium, det fungerar som en slags kuvös för växtembryot. Det vete som användes var tetraploid och rågen diploid. Avkomman efter korsningen blev därför triploid. I en triploid växt fungerar inte meiosen som den ska och växten blir steril. För att rågvetet skulle bli fertilt användes colchicin så att slutprodukten blev hexaploid.
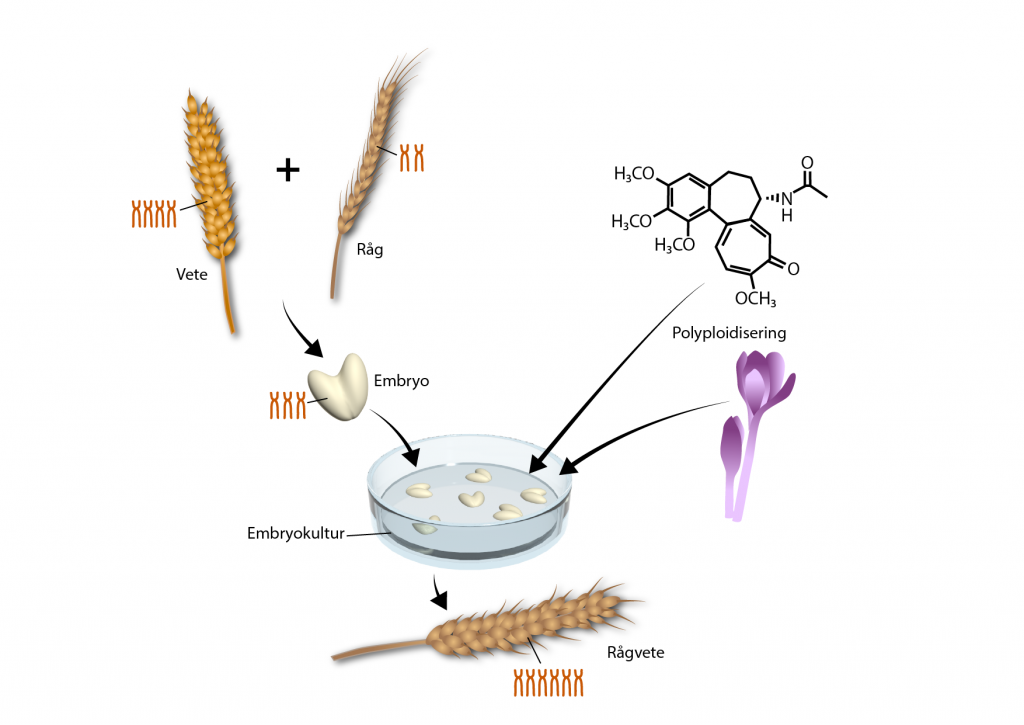
I grödor som råg och vete skördar man frön och grödan måste därför vara fertil. Annars bildas det inga frön. I andra fall är det önskvärt att växten är steril och inte producerar några frön. Exempel på sterila triploida grödor är vattenmelon utan frön (kärnor) och banan. De svarta prickarna i en banans inre är början till frön.

Enligt GMO- lagstiftningen leder inte polyploidisering till en GMO.
På sidan om Bladmögelresistent potatis kan du läsa mer om hur tekniken har använts.
Cell-och vävnadsodling
Cell-och vävnadsodling är en samling tekniker som används för att föröka upp växter eller odla till exempel celler, vävnader eller organ. Det sker under sterila förhållanden på ett näringsmedium. Under de rätta förhållandena kan i princip alla celler i en växt utvecklas till en ny planta med blad, rötter och blommor. Man säger att växtceller är totipotenta.

Mikropropagering
Mikropropagering innebär att en viss planta förökas upp på ett väl definierat näringsmedium. Tekniken används bland annat på växter som inte producerar några livsdugliga frön, till exempel banan, kärnfria vindruvor och många citrusfrukter. Äpplen producerar visserligen frön, men DNA:t i fruktköttet och i kärnorna är inte detsamma. Om frön från till exempel äppelsorten Åkerö får gro blir resultatet inte ett träd med Åkerö-äpplen. Fruktköttet får nämligen sitt DNA från Åkerö, men de frön som bildas i Åkerö-äpplet är en kombination av DNA från Åkerö och den äppelsort som pollinerat.
Meristemodling
En växts tillväxtzoner kallas meristem och består av en grupp celler som kan sägas motsvara människas stamceller. Cellerna i meristemen delar sig och utvecklas till specialiserade celler i växtens olika organ. Vanligtvis finns det inte några virus i meristem-vävnaden. Vill man därför ta fram virusfritt material dissekeras meristem fram och odlas på näringsmedium. Med meristemen som bas utvecklas sedan plantor som är virusfria.
Somatisk hybridisering
Vid somatisk hybridisering, eller protoplastfusion, fuseras somatiska celler. Växtceller omges av en cellvägg som består av cellulosa, hemicellulosa och pektin. För att cellerna ska kunna smälta samman avlägsnas cellväggen med hjälp av enzymer. Kvar blir det plasmamembran som omsluter cellen, en protoplast. När protoplasterna smält samman odlas de på näringsmedium med olika sammansättningar allt eftersom en planta utvecklas. Först delar sig de fuserade protoplasterna och bildar en odifferentierad vävnad, en kallus. För att det ska bildas skott från celler i kallusen måste sammansättningen av växthormoner i mediet ändras. Slutligen rotas skotten på ett medium särskilt anpassat för rotutveckling.

Om de arter vars protoplaster fuseras kan utbyta genetiskt material genom konventionella förädlingsmetoder blir resultaten inte en GMO. Om de däremot inte kan utbyta genetiskt material med konventionella metoder är den somatiska hybriden en GMO som ska regleras.
Embryokultur
När olika växtarter korsas kan det vara svårt för embryot att utvecklas. Om så är fallet kan embryot dissekeras fram och odlas på ett näringsmedium som en typ av kuvös. Där kan ett växtembryo utvecklas till en hel planta.
Genteknisk förädling
Med något enstaka undantag har de genetiskt modifierade grödor som odlas idag förädlats via klassisk genetisk modifiering. Det innebär att en ny DNA-sekvens integrerats i en växts genom. Det finns två viktiga skillnader mellan genetisk modifiering och förädling via korsning:
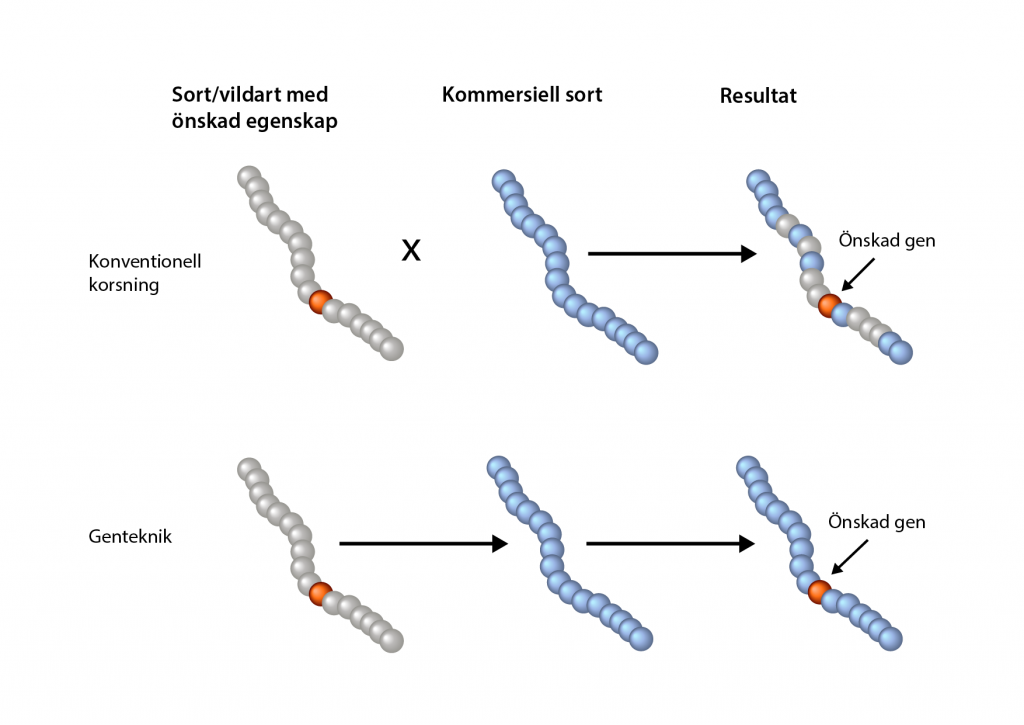
- Vid genetisk modifiering kan enstaka egenskaper överföras från en växt till en annan. En DNA-sekvens, till exempel en gen, isoleras och inga andra gener följer med när DNA-sekvensen förs in i en grödas genom. Vid korsningsförädling blandas DNA:t från de två plantor som korsats och även oönskade gener dyker upp i avkomman.
- Vid genetisk modifiering kan DNA-sekvenser från vilken organism som helst användas. En växt kan till exempel modifieras med en bakteriegen, men en potatis som modifieras med en potatisgen är också en GMO. Anledningen till att det är möjligt är att alla levande varelser är uppbyggda på samma sätt. DNA:t i en människa, en bakterie och en tussilago innehåller samma typ av byggstenar, nukleotider.
Används traditionell mutagenes vid förädlingen skapas slumpmässiga mutationer över hela växtens genom. En gensax som CRISPR/Cas9 kan riktas in mot den plats i genomet som man vill mutera. En gröda som förädlats via traditionell mutagenes regleras inte som en GMO. Riktade mutationer med hjälp av en gensax leder däremot till en växt som regleras på precis samma sätt som en klassisk GMO.
Vävnadsodling vid genetisk modifiering och genomredigering
Vävnadsodling är viktigt vid genetisk modifiering och genomredigering. En växt kan modifieras med hjälp av till exempel bakterien Agrobacterium tumefasciens. Vävnadsbitar från till exempel blad läggs i en lösning av bakterien som för in genen av intresse i några celler i snittytan. När bakterierna gjort sitt jobb odlas bladbitarna på ett näringsmedium under sterila förhållanden. Oftast bildas det först en kallus och sedan ett skott som rotas.
När den svenska amylopektinpotatisen togs fram isolerades först protoplaster. Protoplasterna tog upp CRISPR/Cas9-komplexet som redigerade potatisens arvsmassa. Protoplasterna fick sedan växa och utvecklas till genomredigerade potatisplantor.
Uppdaterad november 2020.