Växtförädling är grundläggande för det mesta vi odlar
Modern växtförädling är viktig för:
- livsmedelsförsörjning och livsmedelssäkerhet. Förädlingsmål är ökad avkastning, ökat näringsinnehåll, minskat matsvinn.
- klimatanpassning av jordbruket. Förädling för ökad tålighet mot torka, värme, salt, och motståndskraft mot sjukdomar och skadedjur som ökar med ett förändrat klimat.
- minskad miljöpåverkan och effektivare resursanvändning. Förädlingsmål är effektivare vatten- och kväve-användning, motståndskraft mot patogener och insektsangrepp som kräver mindre mängd bekämpningsmedel.
- utveckling av växtbaserade råvaror inom bioekonomi och industri. Förädling sker till exempel för ändrat stärkelseinnehåll och vedkvalitet.
Genetisk variation en förutsättning för förädling
För att förädlaren ska kunna ta fram nya sorter med förbättrade egenskaper, måste genvarianter som ger upphov till de önskade egenskaperna finnas tillgängliga hos de växter som förädlaren har tillgång till. Olika varianter av en och samma gen kan ge olika egenskaper. En vetesort kan till exempel ha en genvariant i sitt genom som ger den motståndskraft mot en svampsjukdom, en annan vetesort kan ha en variant av en annan gen som gör den motståndskraftig mot ett visst virus. Förädlaren skulle i det här fallet kunna kombinera de två gynnsamma genvarianterna via att korsning av de två sorterna.
Genetiskt variation, och olika genvarianter, orsakas av att det uppstår förändringar, mutationer, i DNA. Mutationer kan uppstå spontant och ge positiva, negativa eller neutrala effekter och påverkan på växtens förmågan att föröka sig. Ibland är mutation som negativ för en vild växt men positiv ur perspektivet att vi människor vill odla den. Genetisk variation är en förutsättning för evolution av nya arter, men också för förädling av nya sorter.
Samtidigt som genetisk variation är grundläggande för förädling strävar förädlaren efter enhetlighet inom en sort. När lantbrukaren köper frö förväntar han eller hon sig att alla plantor som sås blommar samtidigt, blir ungefär lika höga och mognar och kan skördas samtidigt. För att det ska vara möjligt krävs att det inom en sort är en låg genetisk variation, att växterna inom en sort är genetiskt homogena.
För förädlare är det därför viktigt att ha tillgång till många olika sorter av en art, gärna också lantraser och vilda släktingar, för att så stor genetisk variation och därmed så många olika egenskaper som möjligt ska ingå i förädlingspopulationen. Genbanker som samlar in och bevarar frön är därför viktiga för att så mycket genetisk variation som möjligt ska bevaras för framtiden.
Domesticering – från vilda växter till odlade
Ända sedan vi människor för omkring tio tusen år sedan blev bofasta och började odla växter och hålla lantbruksdjur har vi förändrat sådana växter och djur genetiskt. De växter som hade egenskaper som gav oss fördelar förökades med frön, sticklingar eller ympar. Över många generationer av odling blev de vilda växterna mer och mer anpassade efter människans behov. En evolution driven av ett mänskligt snarare än ett naturligt urval ägde rum. Man säger att både växter och djur domesticerades.
Domesticeringen innebar ofta stora förändringar av de växter människan samlade in. Från att ha varit anpassade till sin naturliga livsmiljö anpassades växterna till människans odlingar. Till exempel selekterades gräs med frön som hålls kvar på plantan när de mognar, och därför enklare kan skördas.
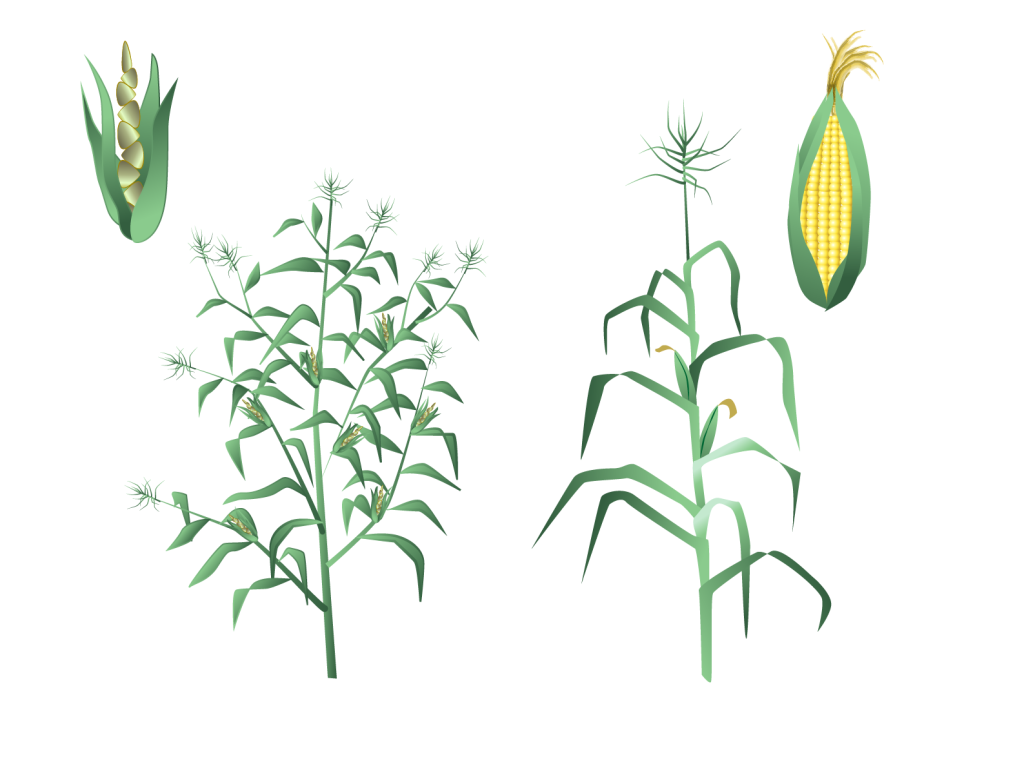
Det här kan man tydligt se i skillnaden mellan den odlade majsen, där majskornen sitter kvar i majskolven, och den vilda majsen, teosint, som har få frön med hårt skal på glesa ax som när de mognar faller till marken. För teosint är det här en viktig egenskap för att den ska sprida sig effektivt. Det hårda skalet gör att fröna går närmast oskadda förbi mag-tarmkanalen hos det djur som äter fröet och kan därför spridas långt. Egenskaper som passar oss människor är i stället många frön (majskorn), med mjukt skal som sitter fast på axet när de mognat. En stadig stam, snarare än ett förgrenat växtsätt är också en fördel. Så ser dagens majs ut, efter tusentals år av mänskligt urval.
Ett annat exempel är de olika typerna av kål. Blomkål, brysselkål, kålrabbi, vitkål och broccoli är alla samma art och härstammar från den vilda kålen (Brassica oleracea). Att det utvecklas ett blomkålshuvud beror på genetiska förändringar (mutationer) i vildkålens blomningsgener som gör att den har svårigheter att blomma. Den försöker, men bildar i stället ett blomkålshuvud som består av många blomanlag som inte utvecklas vidare. Ur kålens perspektiv är det negativt eftersom den inte kan sprida sina gener om den inte blommar, för människan är det positivt eftersom vi får mer att äta. Den typen av mutationer skulle snabbt ha försvunnit ur vildkålspopulationen om vi människor inte tagit hand om och förökat de växter som bar på mutationerna.

Fram till det sena 1800-talet bestod jordbrukets växter av lantsorter där varje enskild planta hade sin unika genuppsättning. I en sådan åker fanns ännu en stor genetisk variation, men också en stor variation i plantornas egenskaper. Den moderna växtförädlingen tog där sin början via ett aktivt urval där de mest lämpade växterna sparades och förökades. Det gav upphov till mer enhetliga sorter där alla plantor hade i princip en identisk genuppsättning.
Korsningsförädling
Enhetliga sorter med olika goda egenskaper kan kombineras i en och samma sort via korsning. Sådan kombinationsförädling har varit dominerande inom växtförädlingen sedan början av 1900-talet, och är det än i dag. Grundläggande för korsningsförädling är goda kunskaper om artens sexuella förökning – är den självpollinerande, eller måste den pollineras av en annan blomma? Själva korsningen är ofta ett pilligt jobb där förädlaren tar pollen från en sort för att pollinera en annan. I processen måste ofta de omogna ståndarna plockas bort från den mottagande blomman för att undvika självpollinering, och blomman måste skyddas från att besökas av insekter som kan bära med sig oönskat pollen. Efter en korsning, blir avkomman bärare av genvarianter från respektive föräldrasort. Anlagen blandas, och för att få tillbaka en homogen sort krävs många generationer av återkorsningar. I den processen måste förädlaren hålla koll på att goda egenskaper inte förloras och att oönskade egenskaper inte introduceras.
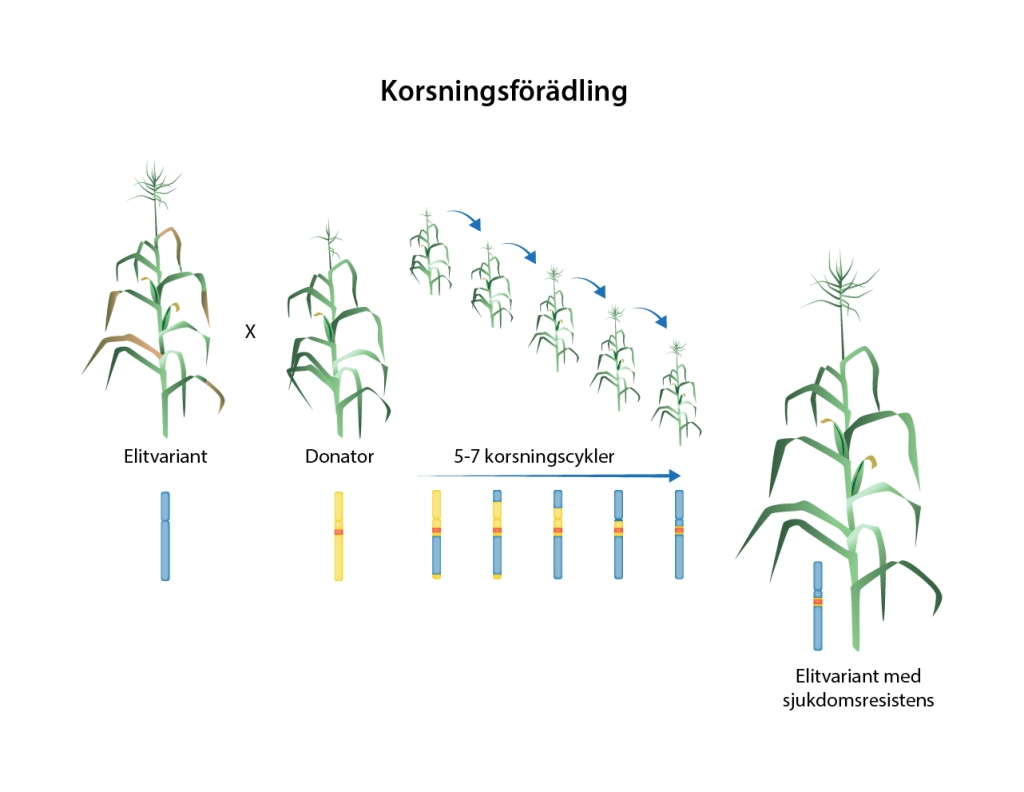
Korsningsförädling är en tidskrävande och kostsam process. En ny sort kan ta 15 år att ta fram. Det betyder att planeringshorisonten för en förädlare är mycket lång och man måste många år i förväg förutse de behov det framtida lantbruket och konsumenterna har och planera förädlingen utifrån det. Med klimatförändringarna är behoven av anpassade sorter än större, planeringen än viktigare och tiden än mer knapp. Modern förädling tar därför till många olika knep för att effektivisera och korta tid och kostnad för att utveckla nya sorter.
Genomisk selektion, accelererad odling, automatiserad fenotypning och artificiell intelligens
Efter en korsning vidtar ett stort arbete med att efter återkorsningar analysera avkommorna för de egenskaper man önskar behålla och sådana man inte vill ha. Många viktiga egenskaper styrs av flera olika gener och därför är förståelse för hur gener samspelar grundläggande för förädlaren.
Idag kan det antal växter som behöver odlas upp och de egenskaper som behöver studeras minskas eftersom det är möjligt att analysera växterna genetiskt på ett tidigt utvecklingsstadium. Med hjälp av avancerad statistik och artificiell intelligens (AI) med maskininlärning kan genotyper kopplas till egenskaper, fenotyper. Det vill säga gener kan kopplas ihop med fysiska egenskaper. Det ger möjlighet att analysera genetiska markörer eller den genomiska sekvensen och bara gå vidare med de växter som med störst sannolikhet har de egenskaper man önskar.
När man söker efter olika egenskaper i förädlingspopulationen eller under förädlingsprocessen måste många olika egenskaper mätas och analyseras hos många växtindivider. Det kallas för fenotypning. Att analysera fenotypiska parametrar som höjd, stamtjocklek, grenars antal och rötters djup och utbredning är mycket arbetsintensivt. Därför har fenotypningen under senare år börjat automatiseras, till exempel genom att växter i växthus på ett löpande band regelbundet avbildas från olika vinkar, rotsystemet studeras med magnetkamera eller växter i fält skannas med hjälp av drönare. Analyserna av de enorma mängden parametrar som kan fås fram stöttas av AI.
Att få en växt att genomgå flera generationer på ett år snabbar också på förädlingsarbetet avsevärt. Det är möjligt genom att modifiera odlingsförhållanden, som dagslängd och temperatur, och därigenom lura växterna att mogna och blomma tidigare. Sådan accelererad odling kallas ”speed-breeding” på engelska, och kan spara mycket tid.
Digitalisering och avancerad dataanalys används i allt större utsträckning i växtförädling. AI-baserade modeller kan analysera stora genetiska och fenotypiska dataset, förbättra prediktioner av växters beteenden i olika miljöer och optimera korsningsstrategier och selektionsbeslut.
Hybridförädling
En specialvariant av korsningsförädling utgörs av hybridförädling. Då korsas plantor från så kallade rena linjer. Rena linjer fås genom att de återkorsats under flera generationer, och är homozygota, det vill säga har två likadana genvarianter av de flesta av sina gener. De plantor som korsas ska komma från rena linjer som genetiskt är så olika varandra som möjligt. Det gör att den första generationer, som kallas F1, från en korsning mellan rena linjer blir starkt heterozygota hybridplantor. De har alltså fått olika genvarianter från sina föräldrar för de flesta gener. Sådana hybridplantor blir påtagligt mer produktiva än sina föräldraplantor. Den effekten kallas hybridvigör eller heterosiseffekten. Den effekten är så pass stor att korsningar för att ta fram F1-hybrid-frön görs för många olika grödor, till exempel majs och sockerbeta, även om det är arbetsintensivt att göra korsningarna och utsädet därmed blir dyrare.
Heterosiseffekten kvarstår dock bara i en generation. Eftersom en F1-hybridplanta är starkt heterozygot kommer olika anlag att i nästa generation dela upp sig och avkommorna till en F1-hybrid blir sinsemellan väldigt olika genetiskt. Mycket forskning ägnas åt att försöka få till asexuell förökning via frön. Det skulle göra det möjligt att fröföröka F1-hybrider och få stabil och enhetlig avkomma som uppvisar heterosiseffekt över många generationer.
Polyploidisering – kromosomfördubbling
Under tidigt 1930-tal upptäcktes att polyploidi, kromosomfördubbling, spelat en viktig roll i växternas evolution och att många viktiga grödor, som potatis och vete var polyploida. Intresset väcktes då för artificiellt framtagna polyploider, eftersom det kan ge större frukter, frön eller blommor, kraftigare tillväxt, bättre kvalitet, sterilitet eller möjlighet att korsa olika arter.
För att inducera polyploidi används det toxiska ämnet colchicin som fås från växten tidlösa. Colchicin påverkar celldelningen genom att hämma bildandet av fungerande mikrotubuli. Uppgiften som mikrotubuli har under celldelningen är att se till att de två celler som blir resultatet av en delning får lika många kromosomer.
Artificell polyploidisering används ibland för att göra det möjligt att korsa två olika arter där avkomman får ett udda antal kromosomuppsättningar, till exempel tre, vilket gör den steril. En fördubbling av kromosomantalet ger då åter ett jämt antal kromosomeruppsättningar (sex i det fallet), och en fertil växt. Ett exempel där polyploidisering använtd på detta sätt är när rågvete, som är resultatet av en korsning mellan råg och vete, togs fram.
I andra fall är det önskvärt att växten är steril och inte producerar några frön. Exempel på sterila triploida grödor är vattenmelon utan frön (kärnor) och banan. De svarta prickarna i en banans inre är början till frön. Nya bananplantor fås genom vegetativ förökning, men för att få en kärnfri, triploid vattenmelon görs korsningar mellan en diploid och en kromosomfördubblad, tetraploid, förälder.

Mutationsförädling – ökar den genetiska variationen
En stor utmaning inom växtförädling är att den genetiska variationen inom en art begränsar vad som är möjligt att åstadkomma med hjälp av korsningsförädling. Genvarianter måste ju finnas i förädlingspopulationen som ger upphov till de egenskaper man önskar introducera i en sort. För att öka den genetiska variationen kan man ta till olika tekniker, både sådana som räknas in i teknikarsenalen för konventionell förädling och sådana som bygger på genteknik.
Nya mutationer kan introduceras i en sort som redan har olika goda egenskaper, men där man önskar ytterligare någon specifik egenskap, till exempel motståndskraft mot en sjukdom. Det här kan göras med mutationsförädling, vilket är en metod med en hundraårig historia. Under 1920-talet upptäckte forskare att nya egenskaper kunde uppkomma hos växter om utsattes för strålning. På 1940-talet började även kemiska ämnen att användas. Både strålning och vissa kemiska ämnen orsakar förändringar i DNA och ger därför en ökad genetisk variation. Strålning ger ofta upphov till deletioner av eller omarrangemang av DNA-sekvensen, medan mutagena ämnen leder till kemisk förändring av vissa av baserna i DNA vilket ger mindre punktmutationer.
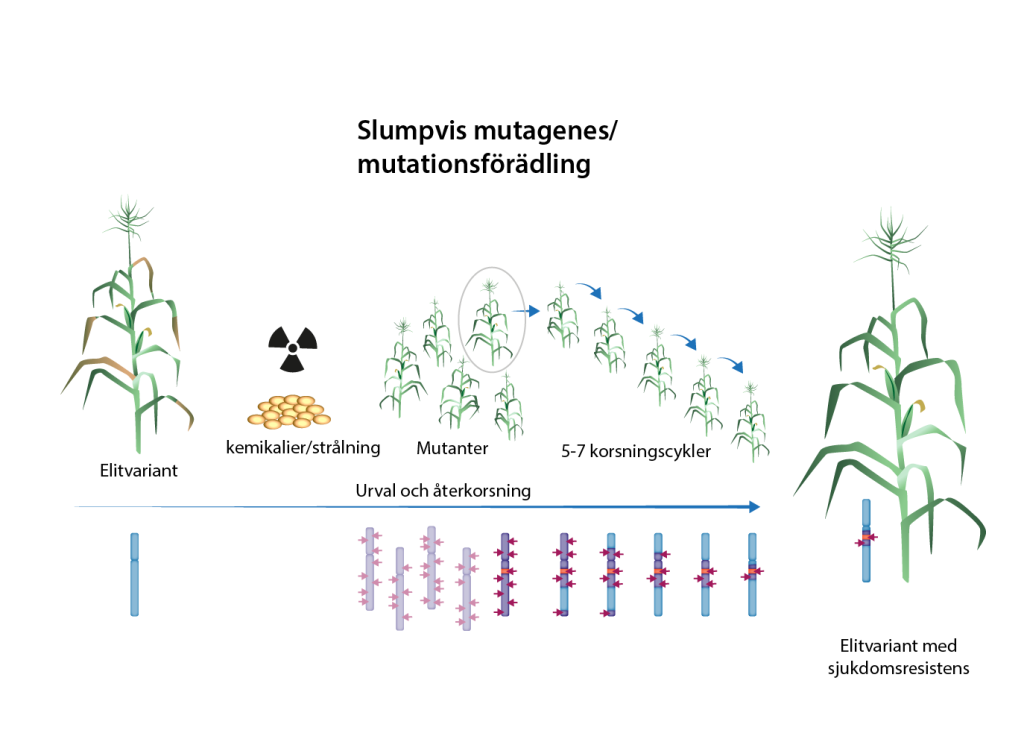
Illustration och copyright: Gunilla Elam
Behandling av växtmaterial med strålning eller mutagena kemikalier ger upphov till tusentals mutationer slumpmässigt utspridda över växternas genom. De flesta mutationer ger inte upphov till förändringar i egenskaper. Man måste därför analysera tusentals plantor för att finna någon med användbara egenskaper.
I och med att så många mutationer introduceras måste ändå återkorsningar till den ursprungliga sorten göras för att bli av med de flesta icke-önskade mutationerna.
Mutationsförädling var särskilt populärt under mitten av 1900-talet, inte minst i Sverige. Under 1980-talet började intresset för mutationsförädling att avta, men vid millenniumskiftet introducerades en ny metod, TILLING (Targeting Induced Local Lesions in Genomes) som gjorde det möjligt att identifiera mutationer som inträffat i en specifik gen. Om man då visste i vilken gen man önskade mutationer kunde man inducera slumpmässiga mutationer, och sedan använda molekylära metoder för att, med lite tur, identifiera en växt som fått mutationer i den gen man önskade.
Idag används oftast det mutagena ämnet etylmetansulfonat (EMS) som ger upphov till slumpmässigt spridda punktmutationer i genomet. Det sker eftersom EMS kemiskt förändrar nukleotiden guanin (G). Det leder till att vissa G-C-baspar i stället blir till A-T-baspar i DNA. Ofta används EMS ihop med en TILLING-teknik kopplad till sekvensering.
Genomredigering – att rikta mutationer till specifika gener
Med tekniker för genomredigering är det möjligt att introducera mutationer på specifika ställen i genomet, till exempel i en specifik gen.
Genomredigering kallas också genredigering och de molekylära verktyg som används kallas populärt för gensaxar. De första teknikerna för genomredigering utvecklades under 1990-talet, men det var med CRISPR/Cas9 som genomredigering fick sitt stora genomslag. Första gången CRISPR/Cas9 användes i växter var år 2013. CRISPR/Cas9, liksom andra tekniker för genomredigering, bygger på enzymer (av gruppen nukleaser) som klipper itu DNA-spiralen på mycket specifika och förutbestämda ställen. I CRISPR/Cas9 är nukleaset som klipper Cas9. Brottet på DNA-spiralen upptäcks av cellens reparationssystem som lagar skadan. Lagningen blir inte exakt, några få baspar kan försvinna eller läggas till och det uppstår en mutation. Det som gör CRISPR/Cas9-tekniken speciellt användbar och användarvänlig, är att Cas9 styrs till det specifika stället (sin målsekvens) med hjälp av ett guide-RNA. Sådana är relativt enkla att designa utifrån den DNA-sekvens man vill att mutationen ska riktas till. Det gör tekniken flexibel, och tillgänglig för många.
Idag när genomet hos många växter sekvensbestämts kan man på förhand välja ut den plats i genomet där nukleaset ska klippa itu DNA-spiralen. Därmed kan man välja var i genomet mutationen ska induceras så att till exempel proteinproduktionen från en viss gen hämmas och grödan får en ny egenskap. I och med att ett guide-RNA ger en hög specificitet är risken för att mutationer ska induceras även på andra ställen låg. Det gör att återkorsningar för att bli av med oönskade mutationer inte är nödvändiga, på samma sätt som det är i traditionell mutationsförädling.
Även om CRISPR/Cas9 ibland gör misstag och orsakar oavsiktliga förändringar (så kallade off-targets) så är det inte jämförbart med de tusentals slumpmässigt utspridda mutationer som framkallas inom mutationsförädling med strålning och mutagena ämnen.
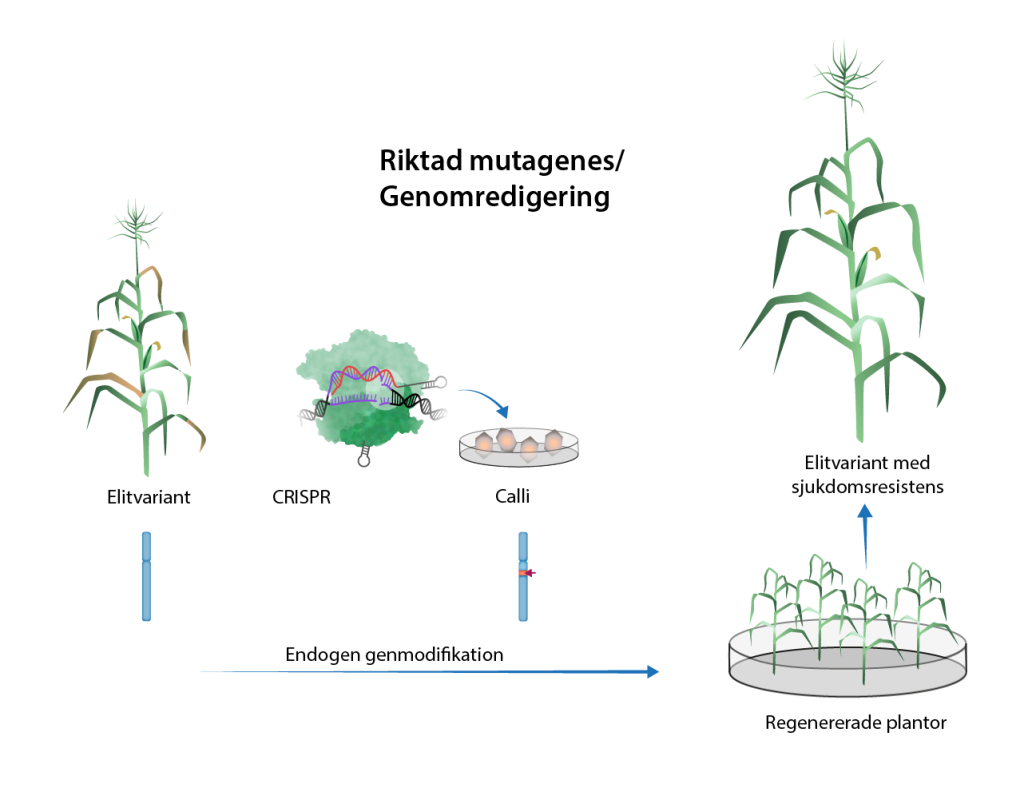
Illustration och copyright: Gunilla Elam
Med genomredigering är det möjligt att ändra gener även i växter som primärt förökas vegetativt. Tekniken används till exempel med framgång för att förädla banan, men även potatis, som visserligen går att korsa men där korsningsförädling är utmanande och mycket tidskrävande bland annat på grund av att potatis är tetraploid.
Det är också möjligt att med hjälp av genomredigering introducera mutationer i flera olika gener samtidigt, även sådana som sitter nära varandra i genomet. Genom korsning är det i princip omöjligt att kombinera mutationer i gener som sitter nära varandra, och då ger den här tekniken nya möjligheter. Till exempel har vete många gener som ger upphov till olika glutenproteiner, och de sitter nära varandra på samma kromosom. Med hjälp av genomredigering har forskare lyckats slå ut ett trettiotal sådana gener vilket resulterat i ett vete med betydligt lägre halt av vissa glutenproteiner. Målet är ett vete med bibehållna bakegenskaper, som personer känsliga mot gluten kan äta.
En utmaning med tekniken är att det krävs en mycket god kunskap om växten och vilka gener som styr en viss egenskap för att man ska kunna göra en eller flera mutationer som får den effekt man eftersträvar. En annan utmaning är att nukleaset måste komma in i växtcellen för att där kunna göra redigeringen. Ibland är det möjligt att skjuta in protein/RNA-komplexet i cellen, men ofta används ett steg där gener som kodar för Cas9-enzymet och guide-RNA:t introduceras i växtens genom. De korsas sedan bort, men i processen går växten då igenom en transgen fas (se nedan).
Transgenteknik – introducera nya gener från en annan organism
Med något enstaka undantag har de genetiskt modifierade grödor som odlas idag förädlats genom att en eller flera nya gener introducerats. På så vis kan specifika egenskaper överföras från en organism till en annan. Det är i princip möjligt att använda gener från vilken organism som helst, eller rent av gener uppbyggda av syntetiskt DNA. Det beror på att DNA är kemiskt uppbyggt på samma sätt hos alla levande organismer, från växter och djur till svampar och bakterier. En vanlig modifiering är så kallade Bt-grödor som fått en gen från en specifik bakterie, Bacillus thuringiensis. I växten avläses genen och ger upphov till ett protein som ger den motståndskraft mot skadegörande insektslarver.
Processen då en gen överförs från en annan obesläktad organism kallas transgenes och en sådan växt kallas transgen.
Det är förstås också möjligt att överföra en gen från en växt inom samma art. Den genen hade då också varit möjlig att överföra via korsning. Men korsning innebär samtidigt att föräldraplantornas alla anlag blandas, vilket ger behov av återkorsningar (se ovan). Att flytta en enstaka gen med hjälp av genteknik gör att återkorsning inte krävs, i och med att den mottagande växtens gener inte i övrigt ändras. Att använda genteknik på det här sättet kallas cisgenes.
Oftast används tekniker som introducerar den nya genen på ett slumpmässigt ställe i genomet. Med CRISPR/Cas-teknik är det möjligt att introducera nya gener på en specifik plats i genomet. På så vis kan en ny gen introduceras på ett ställe i genomet där man vet att den inte påverkar någon annan funktion. Ännu är detta ett ovanligt tillvägagångssätt.
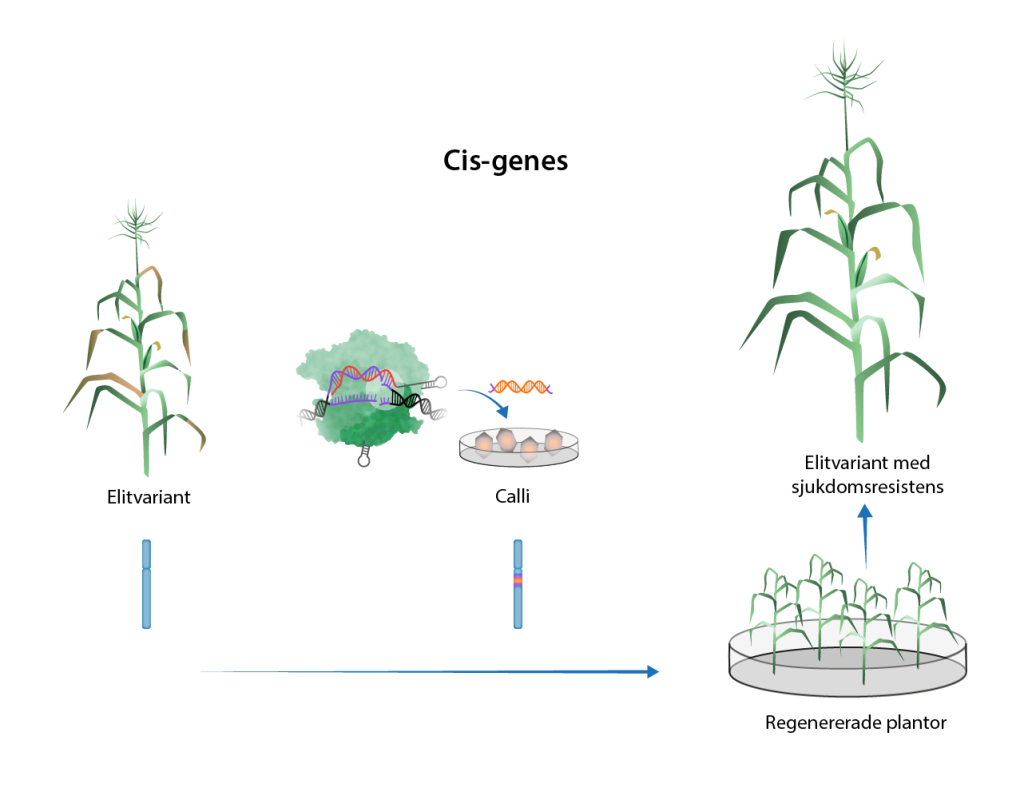
Mer information om hur man använder genteknik i växter finns på sidan om Genetisk modifiering av växter.
Reglering av genetiskt modifierade växter
Enligt EU:s lagstiftning definieras en genetiskt modifierad organism (GMO) som ”en organism, med undantag för människor, i vilken det genetiska materialet har ändrats på ett sådant sätt som inte sker naturligt genom parning eller naturlig rekombination.”
Transgena växter är där med GMO, och ska riskbedömas för att säkerställa att de inte ger negativ påverkan på människors och djurs hälsa eller på miljön.
Som naturlig process räknas korsningsförädling och polyploidisering, men däremot inte mutagenes. Slumpmässig mutagenes med strålning eller mutagena ämnen undantas dock reglering enligt GMO-lagstiftningen, och behöver inte riskbedömas. Anledningen till att dessa växter undantas reglering, är att metoden använts så länge att den anses ha en historia av säker användning.
Riktad mutagenes med CRISPR/Cas9 eller andra liknande så kallade nya genomiska tekniker (NGT), undantas dock inte reglering inom EU. En ny lagstiftning, den så kallade NGT-förordningen, väntas dock godkännas under 2026, och kan i så fall träda i kraft ett par år senare. Den innebär att för växter förädlade med NGT som kan anses ha fått genetiska modifieringar som också skulle ha kunnat uppkomma med konventionella förädlingstekniker inte regleras enligt GMO-lagstiftningen. Omkring 30 andra länder i världen har antagit en liknande lagstiftning för genomredigerade växter där inget nytt DNA introducerats.
Uppdaterad 2026-04-24.